
Research
Research
1. Ovule initiation
Ovule is the precursor of the seed and ovule initiation is the first step of seed formation. Ovule primordium develops into unfertilized seeds after its initiation and following series of developmental processes. In plants characterized by multi-ovulate ovaries, ovule initiation determines the maximal ovule number per flower, thus greatly affecting seed number per fruit and consequently seed yield. Furthermore, as a kind of lateral organ, ovule initiation is also an essential step of plant reproductive development, including cell fate determination, the identity of ovule primordia and boundaries, and ovule protrusion. The regulatory mechanism of ovule initiation is one of the fundamental questions in plant reproductive development. However, ovule initiation remains largely unclear because ovules initiate from placentae which are very tiny and covered by multiple layers of other tissues.
1) The regulatory mechanism of ovule initiation in Arabidopsis
In Arabidopsis thaliana, many ovules protrude from the same placenta, previous studies of ovule initiation focused on a single ovule, little is known about the relationships among individual ovules in the same placenta during ovule initiation. Do all ovules in one placenta initiate at the same time or in a specific order? Whether the initiation of all ovules in a placenta is connected as one event? Are there interactions between the initiation of adjacent ovules? Our findings illustrated older and younger ovules are typically arranged at regular intervals along the placenta, and demonstrated that this phenomenon is caused by asynchronous ovule initiation and similar speed of ovule growth. Besides, the first two batches of ovule initiation didn’t cover the whole number of ovules, we further revealed that there is sporadic new ovule initiation after first two batches. We defined the first two batches of ovule initiation as "early ovule initiation" and the sporadic new ovule initiation as "late ovule initiation". Furthermore, our recent work demonstrated that the dynamic polar localization of PIN1 and the subsequent dynamics of the peaks of auxin signal response are the reasons for the asynchronous ovule initiation. A computational model has been built to mimic the dynamics of auxin signal regulating the asynchronous initiation of ovule primordia. And further results demonstrated that PIN3 and PIN3-mediated auxin response peaks are required for late ovule initiation.
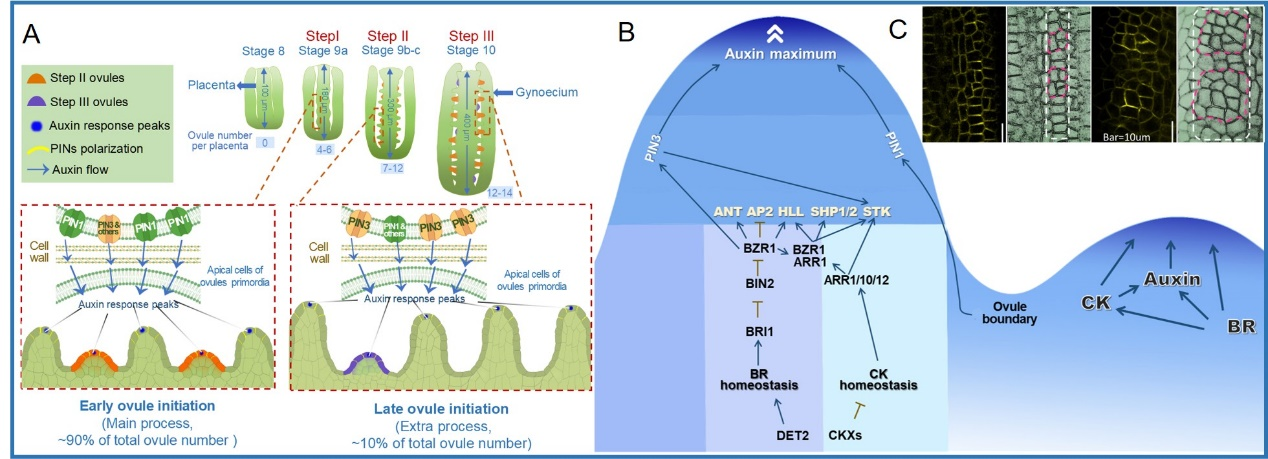
Figure 1. Asynchronous initiation of ovule primordia in Arabidopsis.
2) Signal network of hormonal regulation of ovule initiation and ovule number
Our work demonstrated that plant hormone BR positively regulate ovule initiation, thereby affecting ovule number and seed number per silique, through transcriptional regulating ovule initiation related genes by downstream important transcription factor BZR1, and demonstrated that BR promotes early ovule initiation by enhancing auxin signal, and BR directly regulates the PIN3 expression to promote late ovule initiation. BR and CK crosstalk through the direct interaction of their downstream important transcription factors BZR1 and ARR1, which more effectively induces the expression of ovule initiation related genes to promote ovule initiation and increase ovule number.
2. New mechanism of seed development
Ovule initiation sets the foundation for seed formation. The subsequent processes in ovule, including ovule development, female gametophyte development, male-female interaction, and zygote (embryo and endosperm) development, are also crucial to the final seed yield. Embryo development is the essential part of seed development. Our previous work firstly reported that BR transcriptional regulates the genes related to embryo, endosperm and integument development through BZR1, which affects seed development and regulates seed size and shape.
Our recent work identified that lacking two types of tonoplast proton pumps led to abnormal embryo development through affect vacuole genesis and distribution and polar auxin transport and auxin signal. It is speculated that the tonoplast proton pumps regulate the vacuole-related vesicle trafficking of PIN1, thereby affecting the polar auxin transport and auxin response, and further regulating embryo cell division and pattern formation.
Currently, our preliminary results illustrated that lacking tonoplast proton pumps also affects vacuole formation in embryo sac, thereby influencing the nuclear migration and localization in embryo sac, which leads to irregular development of female gametophyte and seed. Polar auxin transport from sporophyte to gametophyte and the function of BR signal component are also involved in large vacuole formation in embryo sac and affect embryo sac development.
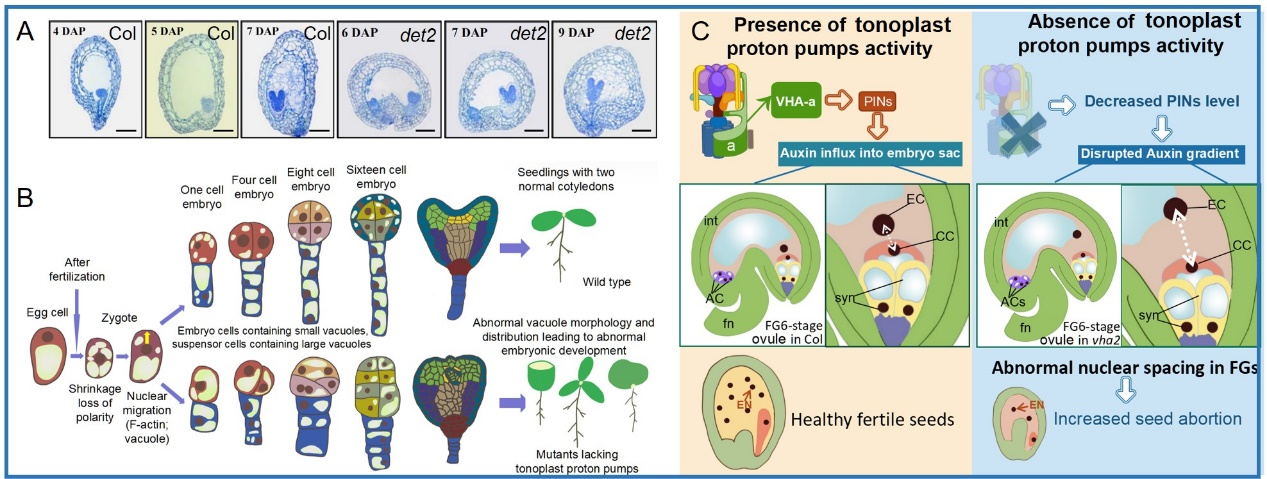
Figure 2. Vacuoles and tonoplast proteins affect female gametophyte and embryo development.
3. BR regulation of plant development and yield-related traits
Plant growth and development have great impact on seed yield, but BR cannot be directly applied to rice production because BR regulates many processes of rice growth and development and induces side effects, such as increasing plant height and lamina joint which causes easy lodging and reduced plant density. To use BR to increase seed yield and avoid side effects, we proposed two strategies to manipulate BR signal to avoid side effects and effectively improve total seed yield. One is to identify new regulators downstream BR signal and participate in the partial process of BR signal regulating plant growth and development, and the other is to modulate BR signal in different organs or different developmental stages. For the first strategy, new rice GATA transcription factors were identified. The total yield per unit area of OsGATA7-cri can be improved by increasing planting density. OsGATA6 positively regulates panicle development and increases grain number without affecting architecture, which has potential application in rice production. According to the second strategy, the reproductive organ specific promoter is used to drive the modified BZR1 coding sequence to directly improve the BZR1 protein accumulation and BR signal in reproductive organs, resulting in improved seed yield in Arabidopsis. In the weak allele of BR-insensitive mutant with sufficient nutrition, the transformation of above construct can lead to the ideal combination of smaller vegetative organs and larger reproductive organs, resulting in dramatic improvement of total seed yield. This system will be introduced into rice in future work.
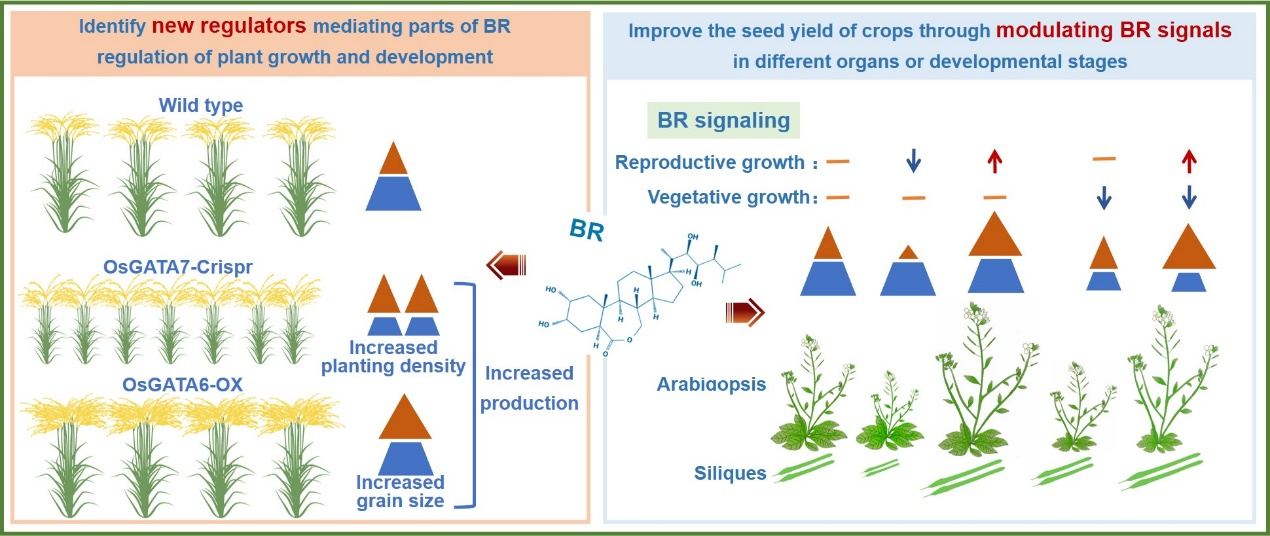
Figure 3. Two strategies manipulating BR signal to effectively improve total seed yield.